Of Six Prehistoric Central Plains Sites
This paper appeared in a slightly modified form in the 1994 Issue of Central Plains Archaeology.
List of Figures
- Rainbow Site, area of maximum sympatry
- Wallace Site, areas of maximum sympatry
- Wallace Site, area of secondary sympatry
- Packer Site, area of maximum sympatry
- Areas of total sympatry from Woodland sites: Rainbow, Wallace, and Packer, excluding range of Microtus Pennsylvanicus
- Hulme Site, area of maximum sympatry
- Palmer Johnson Site, area of maximum sympatry
- Witt Site, area of maximum sympatry
- Areas of total sympatry from Central Plains Tradition sites: Hulme, Palmer Johnson, and Witt, excluding range of Orozymus Palustris
Archaeology, as a holistic science, cannot ignore the biological and climatic environment in which humans have lived. Non-cultural materials such as floral and faunal remains found in an archaeological context can provide clues to the natural environment that ancient peoples inhabited.
Sympatry mapping techniques map the modern regions of habitation for animals found at an archaeological site, to discover what areas the majority of species may be found in today. For example, if five species were discovered at a certain site, and their ranges were mapped over each other, the area where most or all of the species’ modern ranges overlap is the area of maximum sympatry. This mapping can reveal useful information about paleoclimates. Small mammals are especially sensitive to alterations in microclimate, moisture, and temperature, and tend to thrive under very specific conditions. A collection of animal ranges may reflect a narrow climatic band. By finding the area of maximum sympatry today, we can extrapolate the climate of a site several hundred or thousand years ago. For example, in a radical case of sympatry, the site might lie completely outside the area of maximum sympatry. If the area of maximum sympatry is to the north of the site, in terrain that is drier and cooler than the region the site is in today, then archaeologists may consider the site to have been colder and drier at the time it was deposited (Graham and Semken 1987:7–8).
For this study, the author collected faunal data on six Woodland Tradition and Central Plains Tradition sites in Nebraska, Iowa, and Kansas, and applied sympatry mapping techniques to the results. The six sites studied are: the Hulme Site (25HL28), a Central Plains Tradition site in Hall County, Nebraska; the Packer Site (25M9), a Late Woodland Tradition site in Sherman County, Nebraska; the Palmer Johnson Site (25BU37), a Central Plains Tradition site in Butler County, Nebraska; all four horizons of the Rainbow Site (13PM91), Middle to Late Woodland site near Hinton, Iowa ; the Wallace Site (25GO2), a Middle Woodland site in Gosper County, Nebraska; and the Witt Site (14GE600), a Central Plains Tradition site in Central Kansas.
The six sites under consideration in this study were in habitation from AD 250 to AD 1350, following the warm and dry Atlantic Climatic Episode, which ended approximately 5000 years ago. Faunal and pollen analytic studies reveal that the Post-Atlantic Episodes since this time have seen a series of small climatic fluctuations, as climates across the plains and North America became increasingly more like modern climates (Wedel 1986:39–43; Wendland, et al. 1987:466–469). Immediately following the Atlantic Episode was the Sub-Boreal Episode, a period of cool, dry conditions stretching from BC 3000 to BC 950, nearly 1000 years before the earliest sites considered here. Climates became wetter overall in the Sub-Atlantic episode, lasting until approximately AD 300, but returned to warmer and drier conditions for a short period in the Scandic Episode until AD 700. Climates became more moist again for a short period in the Neo-Atlantic, which lasted until AD 1150, but droughtlike conditions began to return to the plains around AD 850, which dominated the continent during the Pacific episode from AD 1150 to AD 1550. The very dry Pacific episode has been cited as a causal factor in the reorganization of subsistence patterns on the plains. Stable agricultural patterns became less prevelant throughout the central plains, shifting to seasonal horticulture in most places, and disappearing altogether in western Nebraska and Kansas and eastern Colorado. Following the Pacific Episode was the Neo-Boreal Episode, from AD 1550 to slightly more than 100 years ago, marked by colder, moister climates in a period known as the “Little Ice Age.” Conditions since this time have generally resembled those of today. (See Table 1 for a summary of Post-Atlantic Episodes).
| Climatic Episode | Summary of Climate | Approx. Dates |
|---|---|---|
| Sub-Boreal | Drier, cooler in northern plains. | BC 3000–950 |
| Sub-Atlantic | Summers wetter, winters possibly stormier. | BC 950–AD 300 |
| Scandic | General return to Atlantic conditions: warmer and drier. | AD 300–700 |
| Neo-Atlantic | Continued warmth, increased moisture. Increasingly droughtlike conditions beginning ca. AD 850, in northeastern plains. | AD 700–1150 |
| Pacific | Droughtlike conditions intensify; seasonally dry air and cooler with decreased preciptation year-round. | AD 1150–1550 |
Table 1: Post-Atlantic climatic episodes, based on pollen and faunal analyses (after Wedel 1986; and Wendland, et al. 1987).
Methods
For this paper, the ranges of certain species found at the sites studied were emphasized over those of others. In general, data concerning large mammals (i.e. larger than rabbit-sized) were not considered for sympatry mapping for three reasons. First, many large animals (such as coyotes and bison) have extensive annual migratory patterns; their ranges can effectively be considered widespread across the North American continent for this reason. In addition, some species (such as bison and antelope) are known to have been actively procured by humans. Their presence in an archaeological context is therefore not necessarily indicative of the local environment. Humans may simply have been bringing in these remains from a distant location. Third, human behavior—especially since European colonization--has had a profound impact on the modern range and numbers of these species. Large predators are especially susceptible to habitat encroachment and bounty hunting, and the great bison hunts conducted by railroad interests in the last century have effectively narrowed bison ranges to nil. Wolves, bison, and, to a lesser extent cougars were probably not uncommon animals to be found across North America 500 years ago, but these animals have all but vanished from most of the Plains in the interim.
The species chosen for this study include any rodent, insectivore, or lagomorph roughly smaller in mass than a rabbit which can be considered a good indicator of paleoclimate.
From the six sites under study, fifteen species were positively identified (with each site averaging seven identified species), and are considered in this study. Specimens where only the family or genus is known, or where their specific classification is unknown, were deemed too uncertain to determine ranges and were not considered. (See Table 2 and Table 3 for a complete list of species and sites studied.)
| Latin Name | Common Name | Family | Sites |
|---|---|---|---|
| Blarina carolinensis | Southern Short-tailed Shrew | Talpidae | 1 |
| Citellus franklinii | Franklin's Ground Squirrel | Sciuridae | 1,2,3,4 |
| Citellus tridecemlineatus | 13-lined Ground Squirrel | Sciuridae | 1,2,4 |
| Cynomys ludovicianus | black-tailed prairie dog | Sciuridae | 1,2,5 |
| Geomys bursarius | plains pocket gopher | Geomydae | 2,3,4,5,6 |
| Microtus ochrogaster | prairie vole | Cricetidae | 1,4,6 |
| Microtus pennsylvanicus | meadow vole | Cricetidae | 3,5 |
| Neotoma floridana | eastern wood rat | Cricetidae | 6 |
| Onychomys leucogaster | northern grasshopper mouse | Cricetidae | 2,3,5,6 |
| Oryzomys palustris | marsh rice rat | Cricetidae | 3,6 |
| Perognathus hispidus | hispid pocket mouse | Heteromyidae | 2,5,6 |
| Scalopus aquaticus | eastern mole | Cricetidae | 5 |
| Sciurius niger | fox squirrel | Sciuridae | 4,5,6 |
| Sylvilagus floridanus | eastern cottontail | Leporidae | 1,2,3,4,6 |
| Synaptomys cooperi | southern bog lemming | Cricetidae | 4 |
Table 2. Species mapped in this study
| Site | Date (AD) | Cultural Affiliation | Climatic Episode |
|---|---|---|---|
| 1. Hulme (25HL28) | 1175–1225 | Central Plains | Pacific |
| 2. Packer (25M9) | 980–1050 | Late Woodland | Neo-Atlantic |
| 3. Palmer Johnson (25BU37) | 1275–1300 | Central Plains | Pacific |
| 4. Rainbow (13PM91) | 250–675 | Mid Late Woodland | Sub-Atlantic to Scandic |
| 5. Wallace (25GO2) | 425–650 | Middle Woodland | Scandic |
| 6. Witt (14GE600) | 1250–1350 | Central Plains | Pacific |
Table 3. Site collections studied.
The modern range of each species represented at a site was mapped on a single transparent overlay, and all the overlays representing the assemblage of small mammals recovered were laid one over another. The area with the greatest overlap of small animal ranges is the area of maximum sympatry. In most cases, this area corresponds to the overlap of all species recovered at a site, but in some cases certain species were found in their modern habitats outside the area of maximum sympatry. These are unusual cases, and have been noted.
Unless otherwise noted, basic data concerning ranges and habitats were obtained from Peterson’s Field Guide to the Mammals, with corroboration from Mammals of the Northern Great Plains.
Middle and Late Woodland Tradition Sites
Data from the Rainbow Site, the Wallace Site, and the Packer Site reveal a picture of the Middle and Late Woodland Tradition (AD 250–1050) climate.
The oldest site, Rainbow, in northwestern Iowa, also has the longest record of habitation. It contains four cultural horizons dating from AD 250 to AD 675. (Falk & Semken 1990: 145–164.) The Rainbow assemblage from all four horizons included: Sylvylagus floridanus, the eastern cottontail; Citellus tridecemlineatus and Citellus franklinii, the thirteen-lined ground squirrel and Franklin's ground squirrel; Sciurius niger, the fox squirrel; Geomys bursarius, the plains pocket gopher; Synaptomys cooperi, the southern bog lemming; and Microtus ochrogaster, the prairie vole. The area of maximum sympatry for all four horizons is fairly large, stretching north into northwestern Minnesota, west to the Nebraska panhandle, south to the Ozarks of Missouri, and along the Mississippi. G. bursarius defines the eastern border of the area of maximum sympatry—generally along the Mississippi River, with a projection into central Illinois and Indiana (Figure 1). All seven of the species are found in this area. The assemblage is dominated by eastern fauna, many of which live in wooded or marshy settings (i.e. Sciurius niger, Sylvilagus floridanus, and Synaptomys cooperi.) The others, however, show a favoritism towards open or meadow settings.
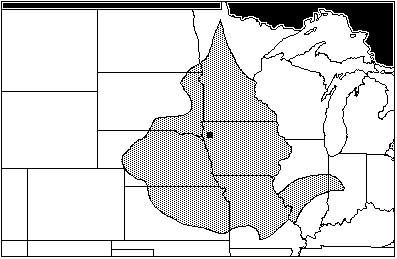
Figure 1. Rainbow Site: Area of Maximum Sympatry (AD 250–675)
The next oldest site of the Woodland affiliation is the Wallace site, in Gosper County, Nebraska, near the Kansas border, and dates from AD 425 to 650. (Bozell & Koch n.d.) The Wallace assemblage includes: Scalopus aquaticus, the eastern mole; Cynomys ludovicianus, the blacktail prairie dog; Sciurius niger; Perognathus hispidus, the hispid pocket mouse; Onychomys leucogaster, the northern grasshopper mouse; Microtus pennsylvanicus, the meadow vole; and Geomys bursarius. There are two small areas of maximum sympatry for the Wallace site, one a sliver of north-central Nebraska running south from the Niobrara river, the other a small wedge in extreme northeastern Colorado (Figure 2). This unusual sympatry is caused by the distribution of Microtus pennsylvanicus, a vole with an affinity for open habitats near water. Today, M. pennsylvanicus is absent from much of Nebraska, including Gosper county. The secondary area of sympatry without M. pennsylvanicus runs along the Niobrara river in the north, south through Nebraska, Kansas and Oklahoma into central Texas (Figure 3). Sciurius niger is the only species of the Wallace assemblage with affinities for woodland habitats; the rest show a definite preference for open, sparsely-vegetated habitat.
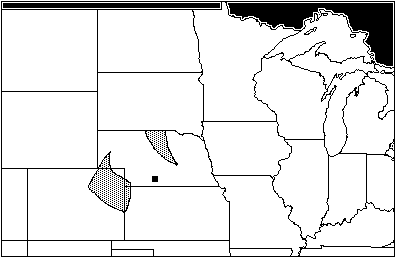
Figure 2: Wallace Site, areas of maximum sympatry (AD 425–650)
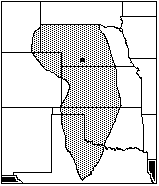
Figure 3: Wallace Site, area of secondary sympatry
The Packer Site, in Sherman County, Nebraska, is the most recent Woodland site, dating to the Great Oasis Phase of the Late Woodland Tradition (ad 980–1050) (Bozell and Rogers 1989:3–36). The Packer assemblage includes: Sylvilagus floridanus, the eastern cottontail; Citellus franklinii; Citellus tridecemlineatus; Cynomys ludovicianus; Geomys bursarius; Perognathus hispidus; and Onychomys leucogaster. The area of maximum sympatry includes all seven of the species, and occupies central Nebraska, extending north into South Dakota, and south into Kansas (Figure 4). All of the species show a marked affinity for low vegetation, short- and long-grass prairie habitats, except for Sylvilagus floridanus. S. floridanus is often considered a woodlands rabbit, but can adapt readily to any low-altitude habitat with adequate ground cover.
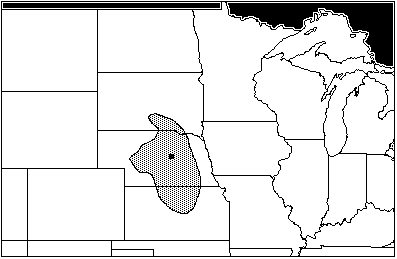
Figure 4: Packer Site, area of maximum sympatry (AD 980–1050)
Taken together, the collected assemblage for the Woodland traditions sites have no area of total maximum sympatry, owing to the unusual modern distribution of Microtus pennsylvanicus. Excluding M. pennsylvanicus leaves an area of sympatry in central Nebraska, extending south into Kansas (Figure 5).
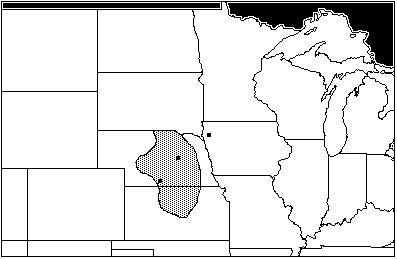
Figure 5: Areas of total sympatry from Woodlands sites: Rainbow, Wallace, and Packer, excluding range of Microtus Pennsylvanicus (AD 2507–1050)
Central Plains Tradition Sites
The Central Plains Tradition existed during the transition from the proposed Neo-Atlantic episode to the Pacific episode. This change is usually marked as being a transformation from warm, moist, Atlantic-dominated climates that made agriculture feasible, to the drier Pacific episode beginning around AD 1150, which may have made seasonal horticulture a more viable subsistence strategy (Bozell 1991: 243–246). Data from the Hulme Site, the Palmer Johnson Site, and the Witt site were considered in evaluating the Central Plains Tradition climate.
The Hulme Site, in Hall County, Nebraska, dates to ad 1175–1225. (Bozell 1991:229–253). Its assemblage contains: Blarina carolinensis, the southern short-tailed shrew; Sylvilagus floridanus; Citellus franklinii; C. tridecemlineatus; Cynomys ludovicianus; Geomys bursarius; and Microtus ochrogaster. The site lies in nearly the center of the area of maximum sympatry, which runs from southern South Dakota through Nebraska and into Kansas (Figure 6). This assemblage represents a variety of habitats, with Plains animals (Citellus tridecemlineatus, Cynomys ludovicianus); mixed-habitat animals (Citellus franklinii); and woodland animals (Blarina carolinensis, Sylvilagus floridanus), although these last three are unparticular animals.
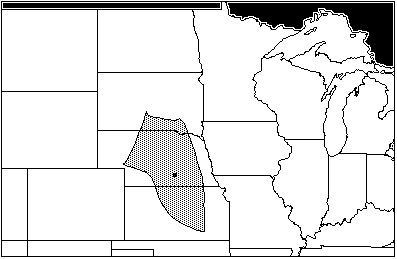
Figure 6: Hulme Site, area of maximum sympatry (AD 1175–1225)
The Palmer Johnson Site, in Butler County, Nebraska, dates to ad 1275–1300. (Ludwickson, et al. 1993). Its assemblage includes: Sylvilagus floridanus; Citellus franklinii; Geomys bursarius; Oryzomys palustris, the marsh rice rat; Onychomys leucogaster; and Microtus pennsylvanicus. The site lies on the southern end of the area of maximum sympatry, which stretches north along the Minnesota/Dakota borders to Canada (Figure 7). The area of maximum sympatry incorporates the ranges of only five of the six species; Oryzomys palustris is found south of the site. O. palustris is the marsh rice rat, a species concentrated mostly in the southeastern U.S., but with a northern limit that may fluctuate hundreds of miles. This may not be a response to climatic change per se, but rather a contraction due to cultural processes. O. palustris prefers seasonally wet habitats, and may have been more plentiful in Nebraska before the draining of floodplain wetlands by Europeans for agriculture in the last century. Alternatively, it has been postulated that rice rats have a preference for maize-cultivated areas, and spread with maize agriculture northwards, only to see a decline in habitat due to the introduction of wheat agriculture, and competition from the introduced R\Rattus norvegicus (Norway rat) (Bardwell 1981:32–38).
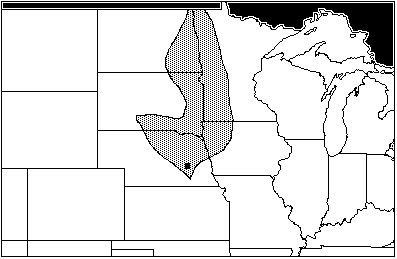
Figure 7: Palmer Johnson Site, area of maximum sympatry (AD 1275–1300)
The Witt Site in north/central Kansas dates to AD 1250- 1350 (Brown 1980: 19- 39). Eight species are represented at the Witt Site: Sylvilagus floridanus; Sciurius niger; Geomys bursarius; Perognathus hispidus; Oryzomys palustris; Onychomys leucogaster; Microtus ochrogaster; and Neotoma floridana, the eastern wood rat. The area of maximum sympatry represents seven of these eight species and covers most of Kansas except for the extreme eastern end, and extends north through Nebraska into South Dakota, west into northeastern Colorado, and south into central Oklahoma (Figure 8). Again, Oryzomys palustris lies outside the range of maximum sympatry, to the east and south.
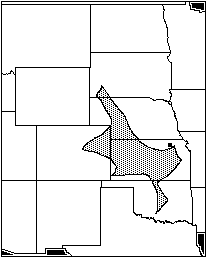
Figure 8: Witt Site, area of maximum sympatry (AD 1250–1350)
The collected sympatry data for all species at Central Plains Tradition sites yields no area of total maximum sympatry, owing to O. palustris. Excluding O. palustris from the data set reveals an area of sympatry covering central Nebraska, extending south into Kansas (Figure 9). This is very similar to the area of maximum sympatry for Woodlands sites, which has implications for the severity of climatic change between the Neo-Atlantic and Pacific episodes ("Conclusions," below).
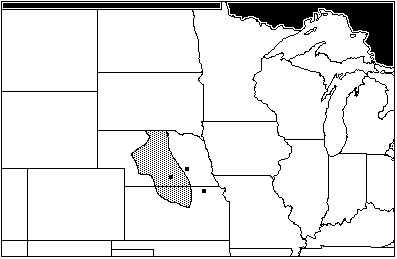
Figure 9: Areas of total sympatry from Central Plains Tradition sites: Hulme, Palmer Johnson, and Witt, excluding range of Orozymus palustris (AD 1175–1350)
Caveats and Cautions
The sympatric-mapping method is an imprecise tool, at best, for determining paleoclimatic conditions. It should be used as only one tool in the paleontological and zooarchaeological toolbox for figuring out past climates, since several factors may skew sympatry maps. First, some species, even the relatively small species considered here, are highly mobile or have wildly fluctuating ranges. The marsh rice rat (Oryzomys palustris), for example, has been a constant thorn in the researcher's side, as the northern extent of its range has been known to fluctuate several hundred miles, possibly due to cultural processes largely unrelated to climatic variation (“Central Plains Tradition Sites,” above).
Second, the level to which these small animals have been affected by human habitation is poorly-understood. These are small and unglamorous creatures, and subsequently not the subject of much study. A few, such as the blacktail prairie dog (Cynomys ludovicianus) and the plains pocket gopher (Geomys bursarius) have been actively hunted or exterminated as pests in many parts of their original ranges. Some species, such as the fox squirrel (Sciurius niger), have thrived in and near urban and suburban habitats, while others, such as the northern grasshopper mouse (Onychomys leucogaster) have faced serious competition from introduced species like the house mouse (Mus musculus).
Third, it is unknown to what level these animals were exploited by humans for food resources. Ethnohistoric records indicate that the larger animals, such as the blacktail prairie dog (Cynomys ludovicianus), were probably used as food by the Omaha, the Pawnee, the Kansa, and other groups; the eastern cottontail (Sylvilagus floridanus) almost certainly was. That these were procured animals has implications for there presence at an archaeological site: just how far would a band of foragers transport rabbits and prairie dogs for food?
Fourth, it is uncertain to what degree some of these animals are intrusive. Certainly, burrowing species such as the eastern mole (Scalopus aquaticus) and the plains pocket gopher (Geomys bursarius) may have intruded into the cache pits and midden pits that were later excavated, and foraging species, such as the hispid pocket mouse (Perognathus hispidus) are probably also not directly associated with the deposition of the artifacts. The question becomes, then, at what time after deposition did these animals intrude? Certainly, there would be no incentive for a mouse to enter a cache pit for food several years after it was abandoned; the food would have decayed by this time. It is entirely likely that most of these animals entered the present-day site within a few decades after deposition, since it is during this time that soil was softest for burrowing, and the proverbial scraps from the humans' tables were still fresh. Moreover, these species, often considered intrusive, may have been used as a food source (Falk and Semken 159–162).
Conclusions
Not surprisingly, the data from this project point to climates in the recent past as being not so different from those of the day. If one dismisses the absence of Microtus pennsylvanicus and Oryzomys palustris from the areas of maximum sympatry as flukes, then the sites all fall within the modern ranges of these animals. Both of these animals have widely-fluctuating ranges, so it is not entirely unlikely that in a few years, they will both be back in the area of maximum sympatry.
Equally as interesting, however, is the comparison of the total sympatric area of all three Woodland Tradition sites, as compared to the total sympatric area for all three Central Plains Tradition sites. Although these maps represent spans of centuries on either side of the transition from the Neo-Atlantic to the Pacific episodes, they are nearly identical, implying that small fauna were not generally affected by the climatic shift.
This in and of itself, however, evokes intriguing possibilities. The Middle Woodland to Central Plains Traditions period encompasses an important era in Plains prehistory: namely, the spread of agriculture through the Plains after the Atlantic Episode, followed by its abandonment in the dry Pacific Episode. That humans may have been evidencing adaptations to climatic changes that left small mammal unaffected says something of both the resilience of humans and the fragility of cultural systems. As a species, we may be more successful and adaptive than any of these fifteen small mammals, but our cultural systems may not be. Very slight changes in microclimate may force ever-widening rings of change in economic strategies, affecting social structure, information networks, and technology.
Modern humans have forgotten how intricately their cultural systems are tied to the natural environment. If climatic shifts so slight as to leave mice and voles unaffected caused a wave a change across the prehistoric Plains, how could global warming, species and ozone loss, and depletion of resources affect modern culture?
Areas for Further Research
Before sympatric-mapping studies can be used as an incontestably reliable tool in paleoclimate studies, two other areas should be explored. As archaeologists become more holistic in approaching entire human ecosystems, it is also necessary to study surrounding, non-human ecosystems. The two interact intimately with one another, and it is imperative that archaeologists work with paleontologists to study the areas of sympatry for animals recovered at non-cultural sites. This will help provide framework for understanding the environment with which humans interacted, without the bias of human intervention into the environment. It will become, essentially, a paleoclimatic control for sympatry mapping in an archaeological context.
At the same time, archaeologists should turn their attention to animal ethology, to gain a better understanding in animal behavior. Knowing how animals behave in and around recently-deposited cultural remains will help to clear up questions regarding post-depositional intrusions. If we can understand how small animals come to be found in an archaeological site, then we can reach more accurate conclusions as to what their presence implies.
Acknowledgements
I am grateful to Rob Bozell of the Nebraska State Historical Society for the use of his literature and for his editorial assistance, and to LuAnn Wandsnider of the University of Nebraska for her editorial assistance and tutelage.
Bibliography
- Bardwell, Jennifer
- 1981. The Paleoecological and Social Significance of the Zooarchaeological Remains from Central Plains Tradition Earthlodges of the Glenwood Locality, Mills County, Iowa. Master's Thesis, Dept. of Social Studies, University of Iowa.
- Bozell, John R.
- 1991. Fauna from the Hulme Site and Comments on Central Plains Tradition Subsistence Variability. Plains Anthropologist 36: 229–253
- Bozell, John R. and Amy Koch
- n.d. Summary of identified vertebrates recovered from the Wallace Site, Gosper County, Nebraska. Data obtained from personal contact.
- Bozell, John R. and Mary K. Rogers
- 1989. A Great Oasis Fauna from Central Nebraska. Central Plains Archaeology 1 (1): 3–36
- Brown, Marie E.
- 1980. Cultural Behavior as Reflected in the Vertebrate Faunal Assemblages of Three Smoky Hills Sites. Master's Thesis, Dept. of Anthropology, University of Kansas.
- Burt, William Henry
- 1980. The Peterson Field Guide Series: A Field Guide to the Mammals. New York: Houghton Mifflin
- Falk, Carl R. and Holmes A. Semken, Jr.
- 1990. Vertebrate Paleoecology and Procurement and the Rainbow Site. In Woodland Cultures on the Western Prairies: The Rainbow Site Investigations, edited by David W. Benn. Office of the State Archaeologist, University of Iowa
- Graham, Russell W. and Holmes A. Semken, Jr.
- 1987. Philosophy and Procedures for Paleoenvironmental Studies of Quaternary Mammalian Faunas. In Late Quaternary Mammalian Biogeography and Environments of the Great Plains and Prairies, edited by R.W. Graham, H.A. Semken, Jr. and M.A. Graham, 1–17. Illinois State Museum Scientific Papers 22
- Jones, J. Knox Jr.; David M. Armstrong, Robert S. Hoffman and Clyde Jones
- 1983. Mammals of the Northern Great Plains. Lincoln, Nebraska: University of Nebraska Press
- Ludwickson, John; John R. Bozell, Amy Koch, Mary J. Adair, and Robert E. Warren
- 1993. Palmer Johnson: A Central Plains Tradition settlement in the Upper Blue River Basin, eastern Nebraska. Final Report Prepared by the Highway Archaeology Program, Nebraska State Historical Society for the Nebraska Dept. of Roads
- Wedel, Waldo R.
- 1986. Central Plains Prehistory: Holocene Environments and Culture Change in the Republican River Basin. Lincoln: University of Nebraska Press
- Wendland, Wayne M.; Ann Benn, and Holmes A. Semken, Jr.
- 1987. Evaluation of Climatic Changes on the North American Great Plains Determined from Faunal Evidence In Late Quaternary Mammalian Biogeography and Environments of the Great Plains and Prairies, edited by R.W. Graham, H.A. Semken, Jr. and M.A. Graham, 460–472. Illinois State Museum Scientific Papers 22